Строение черепа птиц
Череп птицы очень близок к черепу пресмыкающегося, от которого отличается большей легкостью и очень объемистой мозговой коробкой, заканчивающейся клювом, а с боков несущей огромные глазницы.
Характерно также расположение затылочного отверстия на дне черепа, а не в задней его стенке. У взрослой птицы все кости черепной коробки срастаются до полного исчезновения швов. В этом отношении птицы сходны с птеродактилями и летучими мышами — т. е. тоже с летающими животными. Причина этого сходства объясняется, по-видимому, потребностью в облегчении черепа, так как сочленение при помощи швов требует известной толщины костей.
У молодой птицы, в частности голубя, можно различить следующие кости черепной коробки. В состав затылочного отдела входят 4 типичные затылочные кости, располагающиеся вокруг затылочного отверстия: непарная основная затылочная, парные боковые затылочные и непарная верхнезатылочная. Как и у рептилий, под затылочным отверстием лежит непарный затылочный мыщелок. В слуховой капсуле, как и у пресмыкающихся, развиваются три ушные кости, которые рано срастаются как между собой, так и с соседними костями: верхнеушная — с верхнезатылочной, заднеушная — с боковой затылочной; переднеушная остается самостоятельной. Основу черепа составляет основная клиновидная кость, расположенная впереди основной затылочной и покрытая сверху покровной основной височной (basitemporale). Спереди с ней срастается парасфеноид, образуя рострум. В состав задней части глазницы входят небольшие парные боковые клиновидные и глазоклиновидные кости. Очень тонкая межглазничная перегородка образована в передней части непарной средней обонятельной костью (mesethmoideum). Крышу черепа образуют парные теменные, лобные, предлобные (очень рано срастающиеся с лобными), носовые и чешуйчатые, а самую переднюю стенку глазницы — слезные, все эти кости, как всегда, покровного происхождения.
Череп молодого голубя (по Брэму). I — сбоку; II — снизу; III — сверху: 1 — основная затылочная кость, 2 — боковая затылочная, 3 — затылочный мыщелок, 4 — большое затылочное отверстие, 5 — верхнезатылочная кость, 6 — ушная, 7 — основная клиновидная, 8 — переднеклиновидная, 9 — крыловисочная, 10 — глазоклиновидная, 11 — межглазничная перегородка, 12 — средняя обонятельная кость, 13 — теменная, 14 — лобная, 15 — носовая, 16 — слезная, 17 — межчелюстная, 18 — верхнечелюстная, 19 — скуловая, 20 — квадратноскуловая, 21 — чешуйчатая, 22 — квадратная, 23 — сошник, 24 — крыловидная, 25 — сочленовная, 26 — зубная, 27 — угловая
Еще интересные статьи по теме:
Череп птиц — Википедия Переиздание // WIKI 2
Череп птиц — часть скелета птиц. Череп отличается большой черепной коробкой, бесшовным соединением многих костей и клювом, который заменил у них челюсти с зубами. Вариации челюстного аппарата птиц, его размеров, форм клюва и языка являются одними из самых разнообразных среди всех систем органов птиц.
Способность верхней челюсти перемещаться вертикально относительно черепной коробки называется кинетичностью черепа. Если гибкая зона лежит у основания надклювья, то говорят о прокинетизме, а если она расположена на уровне носовых ям и верхняя челюсть обладает внутренней подвижностью — о ринхокинетизме. Дромеогнатический, или палеогнатический (Paleognathae), тип черепа, при котором сочленение костного нёба и других костей черепа неподвижно, противопоставляют неогнатическому (Neognathae), в который объединяют схизогнатический, десмогнатический и эгитогнатический типы.
Описание
Череп птиц принадлежит к диапсидному типу[1], он имеет два височных окна (верхнее и боковое), нижнюю височную дугу, образованную скуловой и квадратноскуловой костями, верхняя височная дуга утрачена[2]. Чтобы поместить крупный головной мозг [1], птицам потребовалась большая черепная коробка. Облегчению веса черепа способствовало приобретение клюва вместо челюсти с зубами и бесшовное соединение костей черепа у взрослых птиц[1][3]. Исследования окостенения черепа птиц обычно относятся к черепу птенцов[1].
Череп птиц часто сравнивают с черепом пресмыкающимися, у которых он тоже принадлежит к диапсидному типу, но которым не требуется такая большая черепная коробка и более лёгкий клюв (вместо него у рептилий тяжёлые челюсти с зубами)[1][3]. Подвижность надклювья относительно черепа связана с усовершенствованием челюстного аппарата по сравнению с рептилиями[3].
Среди всех систем органов птиц вариации челюстного аппарата вместе с размерами и формами клюва и языка являются одними из самых разнообразных[3].
Строение
Череп молодого гуся (лат. anser) Череп молодого гуся (лат. anser) — вид сзадиМозговой череп
Висцеральный череп
Основанием для верхней челюсти служат крыша черепа и срединная обонятельная кость[1]. Первичная верхняя челюсть состоит из трёх костей. По сравнению с рептилиями, у птиц отсутствуют поперечная кость и эпиптеригоид, рудимент которого сохранился у некоторых видов[4]. К покровной нёбной кости сзади присоединена крыловидная кость, сочленённая челюстным суставом с квадратной костью, которая может выполнять качательные движения вперёд и назад[1][4]. Она также соединена с чешуйчатой и переднеушной костями черепной коробки и может быть соединена с боковой затылочной[4]. Вторичная верхняя челюсть состоит из преднечелюстной, верхнечелюстной и носовой костей, с доминирующими размерами первой из них
Базиптеригоидные сочленения — скользящие сочленения с боковой поверхностью крыловидной кости, образованные выростами на боковых сторонах клиновидной кости (базиптеригоидными отростками). Они присутствуют в частности у страусоподобных птиц, тинаму, некоторых трубконосых, гусеобразных, сов. У некоторых видов, в частности пингвинов, чаек, чистиковых, они образуются на стадии эмбриона и исчезают по мере взросления птиц[5].
У основания верхней части надклювья нёбная кость и передний фрагмент крыловидной кости (гемиптеригоид) могут истончаться и образовывать соединительную перемычку (синдесмоз) или сустав [4][5]. Нёбно-крыловидный сустав позволяет крыловидной кости поворачиваться в горизонтальной плоскости, а рамке костного нёба и скуловой луги — деформироваться[6]. Сустав скользит вдоль клювовидного отростка парасфеноида[5] и позволяет широкий размах движений нёба и надклювья, чему также способствует утрата большинством птиц ограничивающего базиптеригоидного сочленения[6].
Задний край ноздри образован верхнечелюстным и межчелюстным отростками носовой кости. У птиц различают округлые и щелевидные ноздри. Округлые ноздри (голоринальный тип) образованы неглубоко разделёнными отростками, которые сходятся под широким закруглённым углом перед линией соприкосновения лобной и межчелюстной костей. Щелевидные ноздри (схизоринальный тип) образованы глубоко расщеплёнными отростками, которые сходятся под острым углом и могут заходить за линию соприкосновения лобной и межчелюстной костей[7][6]. Такое деление предложил английский зоолог Алфред Генри Гаррод
Нижняя челюсть птиц (подклювье) включает задний отдел, который состоит из замещающей сочленовной кости, угловой, надугловой и предсочленовной костей, и передний отдел, состоящий из покровных костей, зубной и пластинчатой кости. По сравнению с рептилиями, у птиц отсутствует венечная кость[1]. Ветви нижней челюсти некоторых птиц могут выгибаться наружу, так как разделены на две части гибким участком. Из-за этого глотка может раскрываться значительно сильнее, что позволяет заглатывать крупную добычу[1][5]. Такое строение называется стрептогнатизмом и присутствует в частности у трубконосых, веслоногих, чаек [5].
В среднем ухе у птиц расположено стремя — звукопередающая косточка. Подвижную основу скелета стреловидного языка обеспечивают рожки, сформированные из остатков висцеральных дуг и первой жаберной дуги[1].
Кинетичность черепа
Череп вальдшнепа
Кинетичность, или кинетизм, черепа — это способность верхней челюсти перемещаться вертикально относительно черепной коробки[1][5]. Впервые этот термин предложил датский зоолог Ян Верслюис[en] в 1910 году в отношении стегоцефалов и зауроспид, позднее его распространили на рыб и птиц, однако у птиц некоторые варианты кинетичности невозможны[8]. Если гибкая зона лежит у основания надклювья, то говорят о прокинетизме, а если она расположена на уровне носовых ям и верхняя челюсть обладает внутренней подвижностью — о ринхокинетизме[5][8].
В первом случае движения надклювья и квадратной кости синхронизированы, так как нижняя часть основания надклювья соединена с квадратной костью и скуловой дугой
Во втором, надклювье носит стебельчатый характер, так как основание верхней челюсти как правило прорезано щелевидной ноздрёй[1][6]. Гибкая зона захватывает предчелюстную кость и предчелюстные отростки носовых костей[8]. Ринхокинетизм является более примитивной формой[8], им обладают ржанкообразные, ибисовые и журавлеобразные[1], а также голубеобразные, тинаму и бескилевые. У вальдшнепа гибкая зона располагается в конечной трети надклювья, чтобы схватывать червей он может раскрывать только кончик клюва, полностью вонзённого в землю[8].
Кинетичность черепа помогает амортизировать рабочие усилия клюва в случае ошибок. Жёсткость верхней челюсти достигается за счёт напряжения челюстных мышц, которые вызывают кратковременные нагрузки на череп. Кроме того, кинетичностью обусловлено разнообразие производимым клювом движений, а также величина усилия на конце клюва[1].
Типы черепа
В 1867 году английский зоолог Томас Генри Гексли в работе «О классификации птиц и о таксономическом значении изменений в строении костного нёба в этом классе» (On the classification of birds, and on the taxonomic value of the modification of certain of the cranial bones observable in that Class)[9] на основе особенностей строения и взаимоотношения различных частей черепа летающих птиц определил четыре классических типа костного нёба: дромеогнатический, схизогнатический, десмогнатический, эгитогнатический[6]. Дромеогнатический, или палеогнатический (лат. paleognathae), тип черепа противопоставляют неогнатическому (лат. neognathae), в который иногда объединяют все другие типы[5]. Такое деление предложил английский зоолог Уильям Плейн Пайкрафт[en] в 1900 году, который отнёс к группе с палеогнатическим черепом бескилевых птиц. Дальнейшее определение типов вызвало оживлённую дискуссию среди учёных[6].
Дромеогнатический тип
Самый примитивный тип черепа, при котором сочленение костного нёба и других костей черепа неподвижно. У такого черепа нёбные и крыловидные кости соединены неподвижно, базиптеригоидные сочленения хорошо развиты (обычно отсутствуют у других взрослых птиц), сошник крупный, его передний конец сливается с нёбными отростками костей верхней челюсти, а задний конец расщеплён и соединяется расходящимися концами с нёбными и крыловидными костями[5][9].
Длина костного нёба в дромеогнатическом черепе неизменна, продольные перемещения сильно ограничены[10]. Неподвижное соединение не позволяет манипулировать с кормом, оно способно только для амортизации толчков[6] и простых хватательных движений[9]. Дромеогнатический тип является первым этапом эволюции кинетизма черепа у птиц[6]. У современных птиц он характерен для страусоподобных (бескилевых) птиц и тинаму[9].
Схизогнатический тип
Схизогнатический тип черепа является более древним, чем другие неонёбные, которые скорее всего возникли путём его перестройки[5]. При этом типе отсутствует соединение нёбных отросков правой и левой верхнечелюстных костей, между ними расположена длинная щель[5][6]. Сочленение нёбных и крыловидных костей подвижное, они примыкают к клювовидному отростку, отсутствует соединение сошника, заострённого спереди, с нёбными отростками верхнечелюстных костей[5].
Схизогнатический тип характерен для пингвинов, гагар, трубконосых, куриных, большинства журавлеобразных, ржанкообразных, трогонов, большинства сов, козодоевых, колибри, настоящих дятлов[5].
Десмогнатический тип
При этом типе происходит слияние нёбных отростков правой и левой верхнечелюстных костей[5][6] по средней линии из-за чего надклювье становится более прочным и имеет трубчатую конструкцию[6]. Сошник редуцирован либо очень узкий, иногда соединяющийся задним концом с нёбными костями и клювовидным отростком, а передним — со слившимися нёбными отростками[5].
Десмогнатический тип характерен для веслоногих, голенастых, гусеобразных, дневных хищников, некоторых козодоев, ракшеобразных, якамаров, пуховок и туканов из дятлообразных[5].
Эгитогнатический тип
При этом типе нёбные отростки верхнечелюстных костей тонкие и удлинённые, они подходят под сошник, но не соприкасаются ни с ним, ни друг с другом, у широкого сошника свободный передний конец иногда раздвоён, а задний глубоко расщеплён и охватывает клювовидный отросток[5]. Функционально это схизогнатия[6].
Эгитогнатический тип характерен для трехпёрсток, стрижей, бородаток и медоуказчиков из дятлообразных, почти у всех воробьиных[5].
Морфология черепа и систематика
Гексли в 1867 году применил разделение на типы костного нёба для определения четырёх подотрядов обширной группы Carinatae[en]. В современной систематике этот таксономический уровень соответствует надотряду. Гексли не рассматривал родственные отношения между таксонами одной группы, но отметил низкое положение тинаму (Tinamidae)[10]. Образование костного нёба птиц рассмотрел Пайкрафт в 1900 году, который пытался выяснить пути филогенеза. Он обнаружил сходство строения мускулатуры, покровов и репродуктивной системы у птиц с дромеогнатическим нёбом — страусов, бескрылов, моа, эпиорнисов и тинаму — и объединил их в надотряд Palaeognathae, сгруппировав остальных в Neognathae. Формулировки Пайкрафта были основаны на типе соединения между palatina и pterigoidea (шовное против шаровидного), а не на подвижности сустава, что привело к многочисленным дискуссиям в этом вопросе[10].
В 1948 году МакДауэлл (McDowell) рассмотрел костное нёбо птиц как комбинацию многих мелких признаков. В его работе содержится четыре варианта нёба палеогнат и указание на наличие у неогнат сочленений, отличных от шаровидного. МакДауэлл критиковал концепцию Пайкрафта и заключил, что выделение палеогнатического и неогнатического типа необоснованно. Точка зрения МакДауэлла до сих пор поддерживается многими орнитологами[10].
Опровержение концепции Макдауэлла предоставил Хофер (Hofer) в 1955 году, развивая направление биомеханики, основанное Фершлейсом (Versluys) в 1910 году и продолженное Маринелли (Marinelli) в 1928 и 1936 годы и рядом других учёных. Они рассматривали костное нёбо как часть костно-мышечного механизма, нёбно-крыловидный сустав в котором выполняет функцию передаточного звена, от которого зависит размах, частота и сила движений верхней челюсти. Хофер подтвердил концепцию Пайкрафта в целом и значительно укрепил в частностях, но его исследования долгое время не были интерпретированы систематиками[10].
В 1970 году Юдин, наблюдая за живыми птицами в зоопарках, обнаружил различную технику захвата и заглатывания пищи у палеогнат и неогнат. Страусоподобные птицы после захвата пищи делают резкое движение шеей назад, сообщая пище ускорение вглубь пасти, а потом движение вперёд, устремляя голову навстречу заглатываемому объекту. При этом надклювье и нёбо внешне остаются неподвижными, однако небольшой сдвиг создаёт напряжённость в конструкции максиллярного сегмента. Неогнаты совершают движения головой чтобы поставить пищу в контакт с подвижным нёбом и языком, после чего они втягивают кусок вглубь пасти, совершая синхронные движения надклювья, нёба, языка, и, возможно, верхней гортани[10].
На более низких таксономических уровнях, в пределах отрядов, семейств и родов, исследования костного нёба и кинетичности черепа также дают богатый материал об эволюции птиц[10].
Существует также точка зрения, что деление на палеогнаты и неогнаты связано с уровнем совершенствования, то есть палеогнаты сохраняют более низкий уровень развития, а неогнаты – продвинутое состояние, которое развивалось независимо. При этом ни те, ни другие, не могут считаться монофилитическими таксонами, а палеогнаты и негнаты представляют собой грады (уровни развития), а не клады[11].
Номенклатура
| Русское название | Латынь | Примечание | |
|---|---|---|---|
| Мозговой череп (neurocranium) | |||
| Затылочный отдел | |||
| Верхняя затылочная кость | supraoccipitale | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| Боковая затылочная кость | exoccipitale | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| → затылочный мыщелок | condylus occipitalis | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| → затылочное, или барабанное, крыло | ala tympanica | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| Основная затылочная кость | basioccipitale | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| Переднеушная кость | prooticum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → овальное окно | fenestra ovalis | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Заднеушная кость | opisthoticum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → круглое окно | fenestra rotunda | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Верхнеушная кость | epioticum | ОСЖКП. стр. 42 | |
| Клиновидный отдел | |||
| Основная клиновидная кость | basisphenoideum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → базиптеригоидный отросток | processus basipterygoideus | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Боковая клиновидная кость | laterosphenoideum, также pleurosphenoideum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → заглазничный отросток | processus postorbitalis | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Обонятельный отдел | |||
| Срединная обонятельная кость, или обонятельная кость | mesethomoideum | ПЗП. стр. 75 | ОСЖКП. стр. 44 |
| → межглазничная перегородка | septum interorbitale | ПЗП. стр. 75 | ОСЖКП. стр. 44 |
| → черепно-лицевая щель | fissura craniofacialis | ОСЖКП. стр. 44 | |
| Покровный череп (dermatocranium), также висцеральный череп (splanchnocranium) | |||
| Теменная кость | parietale | ПЗП. стр. 76 | ОСЖКП. стр. 44 |
| → височные гребни | linea temporalis | ОСЖКП. стр. 44 | |
| Лобная кость | frontale | ПЗП. стр. 77 | ОСЖКП. стр. 44 |
| Носовая кость | nasale | ПЗП. стр. 77 | ОСЖКП. стр. 45 |
| → лобный отросток | processus frontalis | ОСЖКП. стр. 45 | |
| → межчелюстной отросток | processus praemaxillaris | ОСЖКП. стр. 45 | |
| → челюстной отросток | processus maxillaris | ОСЖКП. стр. 45 | |
| Чешуйчатая кость | squamosum | ПЗП. стр. 77 | ОСЖКП. стр. 46 |
| → скуловой отросток | processus zygomaticus | ПЗП. стр. 77 | |
| → заднеглазничный отросток | processus orbitalis posterior, или sphenoticus | ОСЖКП. стр. 46 | |
| → височный отросток | processus temporalis | ОСЖКП. стр. 46 | |
| Предлобная кость, или слёзная кость | praefrontale | ПЗП. стр. 77 | ОСЖКП. стр. 46 |
| → надглазничный отросток | processus supraorbitalis | ОСЖКП. стр. 46 | |
| → предглазничный отросток | processus orbitalis anterior | ОСЖКП. стр. 46 | |
| Скуловая кость | jugale | ПЗП. стр. 78 | |
| Квадратно-скуловая кость | quadratojugale | ПЗП. стр. 78 | ОСЖКП. стр. 48 |
| Сошник | vomer | ПЗП. стр. 79 | ОСЖКП. стр. 44 |
| Верхняя челюсть (надклювье) | |||
| Вторичная верхняя челюсть | |||
| Предчелюстная, или межчелюстная, кость | praemaxillare | ПЗП. стр. 80 | ОСЖКП. стр. 47 |
| → лобный отросток | processus frontalis | ОСЖКП. стр. 48 | |
| → нёбный отросток | processus palatinus | ПЗП. стр. 81 | ОСЖКП. стр. 48 |
| → челюстной отросток | processus maxillaris | ОСЖКП. стр. 48 | |
| Верхнечелюстная, или челюстная, кость | maxillare | ПЗП. стр. 81 | ОСЖКП. стр. 48 |
| → нёбный отросток | processus palatinus, или maxillo-palatinum | ПЗП. стр. 81 | ОСЖКП. стр. 48 |
| → скуловой отросток | processus zygomaticus | ОСЖКП. стр. 48 | |
| Вторичная верхняя челюсть | |||
| Нёбная кость | palatinum | ПЗП. стр. 82 | ОСЖКП. стр. 44 |
| → каудолатеральная пластинка | lamina caudolateralis | ПЗП. стр. 82 | |
| Крыловидная кость | pterygoideum | ПЗП. стр. 82 | ОСЖКП. стр. 42 |
| → гемиптериоид | hemipterygoideum | ПЗП. стр. 82 | |
| Квадратная кость | quadratum | ПЗП. стр. 82 | ОСЖКП. стр. 48 |
| → ушной отросток | processus oticus | ПЗП. стр. 82 | ОСЖКП. стр. 48 |
| → глазничный отросток | processus orbitalis | ПЗП. стр. 82 | ОСЖКП. стр. 48 |
| Нижняя челюсть (подклювье) | |||
| Сочленовная кость | articulare | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| → внутренний отросток нижней челюсти | processus internus mandibulae | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| → задний отросток нижней челюсти | processus posterior mandibulae | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Зубная кость | dentale | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Подбородочная косточка | mento-mekkelianum | ОСЖКП. стр. 53 | |
| Пластинчатая кость | spleniale | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Угловая кость | angulare | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| → заднеугловой отросток | processus angularis posterior | ОСЖКП. стр. 53 | |
| Надугловая кость | supraangulare | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Предсочленовная, или покровная сочленовная, кость | praearticulare | ПЗП. стр. 84 | |
| Слуховой столбик, или стремечко | columella auris, или stapes | ПЗП. стр. 84 | |
| Подъязычный аппарат | apparatus hyobrahchialis | ПЗП. стр. 85 | ОСЖКП. стр. 54 |
Легенда:
- ПЗП — Гуртовой Н.Н., Дзержинский Ф.Я. Практическая зоотомия позвоночных. Птицы. Млекопитающие. — Москва: Высшая школа, 1992. — 414 с. — ISBN 5-06-002207-2.
- ОСЖКП — Шульпин Л.М. Орнитология (строение, жизнь и классификация птиц). — Ленинград: Издание Ленинградского государственного университета, 1940. — 555 с.
Примечания
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Дзержинский, Васильев, Малахов, 2013, с. 303—306.
- ↑ Дзержинский, 2005, с. 97—98.
- ↑ 1 2 3 4 5 Коблик, 2001, с. 15—16.
- ↑ 1 2 3 4 5 Дзержинский, 2005, с. 90—92.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Карташев, 1974, Кинетизм черепа и типы строения костного нёба.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Дзержинский, 2005, с. 104—107.
- ↑ 1 2 Карташев, 1974, Типы скелетной основы ноздрей.
- ↑ 1 2 3 4 5 6 Дзержинский, 2005, с. 98—103.
- ↑ 1 2 3 4 Коблик, Чернова, 2017.
- ↑ 1 2 3 4 5 6 7 Юдин К. А. Классические морфологические признаки и современная систематика птиц // Русский орнитологический журнал. — 2009. — Т. 18, вып. 498. — С. 1215—1222.
- ↑ Дзержинский, 2005, с. 21.
Литература
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. Зоология позвоночных. — М.: Академия, 2013. — С. 293-. — 464 с. — ISBN 978-5-7965-7971-4.
- Дзержинский Ф. Я. Сравнительная анатомия позвоночных животных. — 2-е. — М.: Аспект-Пресс, 2005. — 320 с.
- Карташев Н. Н. Систематика птиц. — М.: Высшая школа, 1974. — С. 7—. — 342 с.
- Коблик Е. А. Разнообразие птиц (по материалам экспозиции Зоологического музея МГУ). — М.: Издательство МГУ, 2001. — Т. 1. — С. 15—16. — 384 с. — ISBN 5-211-04072-4.
- Коблик Е. А., Чернова О. Древненёбные птицы. Очерки филогении, систематики, биологии, морфологии и хозяйственного использования. — Litres, 2017. — С. 7—8.
- Baumel J. J. Handbook of Avian Anatomy: Nomina Anatomica Avium. — Cambridge: Nuttall Ornithological Club, 1993. — С. 45—132.
 Эта страница в последний раз была отредактирована 20 мая 2020 в 04:34.
Эта страница в последний раз была отредактирована 20 мая 2020 в 04:34.Птицы: череп
Птицы: череп
Череп птиц похож на череп рептилий и может быть отнесен к диапсидному типу с редуцированной верхней дугой. Череп тропибазальный (глазницы расположены впереди головного мозга), образован тонкими губчатыми костями, границы между которыми отчетливо видны лишь у молодых птиц. Это, видимо, связано с тем, что соединение при помощи швов невозможно из-за небольшой толщины костей. Поэтому череп относительно легок. Своеобразна по сравнению с пресмыкающимися и его форма: резко увеличен объем мозговой коробки, глазницы большие, челюсти лишены зубов (у современных птиц) и формируют клюв. Смещение большого затылочного отверстия и затылочного мыщелка на дно черепа увеличивает подвижность головы относительно шеи и туловища.
Большое затылочное отверстие окружено четырьмя затылочными костями: основной (basioccipitale) ( рис. 43 ), двумя боковыми (occipitale laterale) и верхней (supraoccipitale). Основная и боковые затылочные кости образуют единственный (как и у пресмыкающихся) затылочный мыщелок, сочленяющийся с первым шейным позвонком. Три ушные кости, окружающие слуховую капсулу, сливаются с прилегающими костями и между собой. В полости среднего уха находится лишь одна слуховая косточка – стремечко. Бока и крышу мозговой коробки образуют парные покровные кости: чешуйчатые ( рис. 43; 21 ), теменные, лобные и боковые клиновидные (laterosphenoideum) ( рис. 43, 9 ). Дно черепа образует покровная основная клиновидная кость, которую закрывает покровная основная височная кость (basitemporale), и клювовидный отросток парасфеноида (rostrum parasphenoidei) ( рис. 43, 7, 8 ). У его переднего конца лежит сошник, по краям которого расположены хоаны.
Верхняя часть клюва – надклювье – образована сильно разросшимися и слившимися предчелюстными костями ( рис. 43, 17 ). Гребень клюва, укрепленный носовыми костями, соединяется с лобными костями и передней стенкой глазницы, образованной разросшейся средней обонятельной костью (mesethmoideum). Составляющие лишь заднюю часть надклювья верхнечелюстные кости отростками сливаются с небными костями ( рис. 43, 28 ). К задненаружному краю верхнечелюстной кости прирастает тонкая костная перекладина, состоящая из двух слившихся костей – скуловой и квадратно-скуловой. Это типичная нижняя дуга диапсидного черепа, ограничивающая снизу глазницу и височную яму. Квадратно-скуловая кость сочленяется с квадратной костью, нижний конец которой образует суставную поверхность для сочленения с нижней челюстью, а удлиненный верхний конец суставом прикрепляется к чешуйчатой и переднеушной костям. Небные кости концами налегают на клювовидный отросток парасфеноида и суставом соединяются с парными крыловидными костями, которые в свою очередь суставом связаны с квадратными костями соответствующей стороны.
Такое строение костного неба имеет важное значение для свойственного большинству птиц кинетизма (подвижности) надклювья. При сокращении мышц, соединяющих направленный вперед глазничный отросток квадратной кости со стенкой глазницы, нижний конец квадратной кости смещается вперед и сдвигает как небные и крыловидные кости (их соединение друг с другом может скользить по клювовидному отростку), так и квадратно-скуловые и скуловые. Давление по этим костным мостикам передается на основание надклювья и благодаря перегибу костей в области -переносицы- ( рис. 44, I ) вершина надклювья сдвигается кверху. В зоне перегиба надклювья кости очень тонки, а у некоторых видов (гуси и др.) здесь образуется сустав. При сокращении мышц, соединяющих череп с нижней челюстью, вершина клюва сдвигается книзу. Подвижность костного неба в сочетании со сложно дифференцированными жевательными мышцами обеспечивают разнообразные, тонко дифференцированные движения клюва при схватывании добычи, чистке оперения, постройке гнезд. Вероятно, подвижность шеи и приспособление клюва к многообразным движениям способствовали превращению передних конечностей в крылья, так как замещали некоторые из выполняемых ими второстепенных функций (помощь в захвате пищи, чистка тела и др.).
Нижняя часть клюва – подклювье или нижняя челюсть – образуется слиянием ряда костей, из которых более крупные зубная (см. рис. 43, 26 ), сочленовная и угловая. Челюстной сустав формируют сочленовная и квадратная кости. Движения надклювья и подклювья очень четко координированы благодаря дифференцированной системе жевательных мышц. Подъязычный аппарат состоит из удлиненного тела, поддерживающего основание языка, и длинных рожек. У некоторых птиц, например дятлов, очень длинные рожки огибают весь череп. При сокращении подъязычной мускулатуры рожки скользят по соединительнотканному ложу и язык выдвигается из ротовой полости почти на длину клюва ( рис. 45 ).
Ссылки:
Череп птиц Википедия
Череп птиц — часть скелета птиц. Череп отличается большой черепной коробкой, бесшовным соединением многих костей и клювом, который заменил у них челюсти с зубами. Вариации челюстного аппарата птиц, его размеров, форм клюва и языка являются одними из самых разнообразных среди всех систем органов птиц.
Способность верхней челюсти перемещаться вертикально относительно черепной коробки называется кинетичностью черепа. Если гибкая зона лежит у основания надклювья, то говорят о прокинетизме, а если она расположена на уровне носовых ям и верхняя челюсть обладает внутренней подвижностью — о ринхокинетизме. Дромеогнатический, или палеогнатический (Paleognathae), тип черепа, при котором сочленение костного нёба и других костей черепа неподвижно, противопоставляют неогнатическому (Neognathae), в который объединяют схизогнатический, десмогнатический и эгитогнатический типы.
Описание[ | ]
Череп птиц принадлежит к диапсидному типу[1], он имеет два височных окна (верхнее и боковое), нижнюю височную дугу, образованную скуловой и квадратноскуловой костями, верхняя височная дуга утрачена[2]. Чтобы поместить крупный головной мозг[1], птицам потребовалась большая черепная коробка. Облегчению веса черепа способствовало приобретение клюва вместо челюсти с зубами и бесшовное соединение костей черепа у взрослых птиц[1][3]. Исследования окостенения черепа птиц обычно относятся к черепу птенцов[1].
Череп птиц часто сравнивают с черепом пресмыкающимися, у которых он тоже принадлежит к диапсидному типу, но которым не требуется такая большая черепная коробка и более лёгкий клюв (вместо него у рептилий тяжёлые челюсти с зубами)[1][3]. Подвижность надклювья относительно черепа связана с усовершенствованием челюстного аппарата по сравнению с рептилиями[3].
Среди всех систем органов птиц вариации челюстного аппарата вместе с размерами и формами клюва и языка являются одними из самых разнообразных[3].
Строение[ | ]
Череп молодого гуся (лат. anser) Череп молодого гуся (Анатомия птиц
Содержание статьи
Когда птицы обрели способность летать, их строение претерпело заметные изменения по сравнению с тем, которое было свойственно их предкам – рептилиям. Чтобы вес тела животного был по возможности уменьшен, часть органов стала более компактными, тогда как другие был и вовсе утрачены. Что же касается чешуек, то на их место пришли перья.
Те из тяжелых структур, которые имели жизненно важное значение, были перемещены ближе к центру тела, чтобы улучшить его балансировку. Кроме этого, регулируемость, скорость и эффективность всех физиологических процессов заметно повысилась, что обеспечило ту мощность полета, которая требовалась животному.
Скелет птиц
Для птичьего скелета характеры уникальные жесткость и легкость. Облегчение скелета было достигнуто благодаря тому, что ряд элементов был редуцирован (в первую очередь в конечностях птиц), а также благодаря тому, что внутри некоторых костей появились воздухоносные полости. Жесткость же была обеспечена срастанием ряда структур.
В целях удобства описания, скелет птиц делят на скелет конечностей осевой скелет. Последний включает в себя грудину, ребра, позвоночник и череп, а второй состоит из дуговидных плечевого и тазового пояса c костями прикрепленных к ним задних и передних сводных конечностей.
 Строение скелета птицы.
Строение скелета птицы.Строение черепа у птиц
Для птичьего черепа характерны глазницы огромного размера. Их размер настолько велик, что прилегающая к ним сзади мозговая коробка как бы потеснена глазницами назад.
Очень сильно выступающие вперед кости образуют не имеющую зубов верхнюю и нижнюю челюсти, которые соответствуют надклювью и подклювью. Под нижним краем глазниц и почти вплотную к ним расположены ушные отверстия. В отличие от верхней части челюсти у людей, птичья верхняя челюсть подвижна, благодаря тому, что имеет особое, шарнирное прикрепление к мозговой коробке.
Позвоночник птиц состоит из множества мелких косточек, именуемых позвонками, которые располагаются один за другим, начиная от основания черепа до окончания хвоста. Шейные позвонки обособленны, очень подвижны и их как минимум вдвое больше, чем у большинства млекопитающих, включая людей. Благодаря этому птицы могут очень сильно наклонять голову и поворачивать ее практически в любом направлении.
Позвонки грудного отдела сочленяются с ребрами и в большинстве случаев прочно сращены друг с другом. В тазовой области позвонки слиты в одну длинную кость, называемую сложным крестцом. Для таких птиц характерна необычайно жесткая спина. Оставшиеся, хвостовые позвонки достаточно подвижны, кроме нескольких последних, слитых в единую кость называемую пигостилем. По своей форме они напоминают лемех плуга и являются скелетной опорой для имеющих большую длину рулевых хвостовых перьев.
 Анатомическое строение птиц.
Анатомическое строение птиц.Грудная клетка у птиц
Сердце и легкие птицы снаружи защищены и окружены ребрами и грудными позвонками. Быстролетающим птицам присуща чрезвычайно широкая грудина, разросшаяся в киль. Это обеспечивает эффективное прикрепление главных летательных мышц. В большинстве случаев, чем больше у птицы киль, тем сильнее у нее полет. У птиц, которые совершенно не летают, киль отсутствует.
Связывающий крылья с остевым скелетом плечевой пояс с каждой стороны образован тремя костями, которые расположены наподобие треножника. Одна ножка этой конструкции (воронья кость – коракоид) упирается в грудину птицы, вторая кость, являющаяся лопаткой, лежит на ребрах животного, а третья (ключица) сливается с противоположной ключицей в единую кость называемую «вилочка». Лопатка и коракоид в том месте, где они сходятся, образуют суставную впадину, в которой и поворачивается головка плечевой кости.
 Скелет птиц предельно упрощен и образован легкими и прочными костями.
Скелет птиц предельно упрощен и образован легкими и прочными костями.Строение крыльев у птиц
В общем, кости птичьих крыльев те же, что и кости человеческой руки. Точно так же как и у людей, единственной костью верхнего отдела конечностей является плечевая кость, которая сочленяется в локтевом суставе с двумя костями (локтевой и лучевой) предплечья. Ниже начинается кисть, многие элементы которой, в отличие от их человеческих аналогов, слиты между собой или же вовсе утрачены. В итоге остаются только две кости запястья, одна пряжка (пястно-запястная кость крупного размера) и четыре фаланговых кости, которые соответствуют трем пальцам.
Птичье крыло намного легче, чем конечность любого другого наземного позвоночного, сходного по размерам с птицей. И это объясняется не только тем, что птичья кисть включает в себя меньше элементов. Причина еще и в том, что длинные кости предплечья и плеча птицы являются пустотелыми.
 Строение и виды перьев птиц.
Строение и виды перьев птиц.Причем в плечевой кости находится специфический воздушный мешок, который относится к дыхательной системе. Дополнительное облегчение крылу придает то, что крупные мышцы в нем отсутствуют. Вместо мышц главные движения крыльев контролируются с помощью сухожилий очень развитой мускулатуры грудины.
Отходящие от кисти летательные перья называются первостепенными (большими) маховыми перьями, а те которые прикреплены в районе локтевых костей предплечья, называются второстепенными (малыми) маховыми перьями. Помимо этого разливается еще три пера крыла, которые прикреплены к первому пальцу, а также кроющие перья, которые гладко, наподобие черепицы, налегают на основания маховых перьев.
Что касается тазового пояса птиц, то с каждой стороны тела он состоит из трех костей, слитых между собой. Это подвздошная, лобковая и седалищная кости, причем подвздошная кость сращена со сложным по своей структуре крестцом. Такая сложная конструкция защищает почки снаружи, одновременно обеспечивая прочную связь ног с плечевым скелетом. Там, где три кости относящиеся к тазовому поясу сходятся друг с другом, находится значительная по своей глубине вертлужная впадина. В ней вращается головка бедренной кости.
 Анатомическое строение крыла птиц.
Анатомическое строение крыла птиц.Устройство ног у птиц
Как и у людей, бедренная кость птиц является стержнем верхнего отдела нижних конечностей. В коленном суставе к этой кости причленяется голень. Но если у людей в состав голени входит малая и большая берцовые кости, то у птиц они сращены между собой, а также с одной косточкой предплюсны или с несколькими. Вместе этот элемент называется тибиотарзус. Что же касается малой берцовой кости, то от нее остался заметен только коротенький тонкий рудимент, который прилегает к тибиотарзусу.
Устройство стоп у птиц
Во внутрипредплюсневом (голеностопном) суставе, к тибиотарзусу причленяется стопа, которая состоит из одной длинной кости, костей пальцев и цевки. Последняя образована элементами плюсны, которые сращены между собой, а также несколькими предплюсневыми нижними косточками.
 Анатомическое строение ног птиц.
Анатомическое строение ног птиц.Большинство птиц имеет четыре пальца, каждый из которых причленен к цевке и заканчивается когтем. Первый палец у птиц обращен назад. Остальные пальцы в большинстве случаев направлены вперед. Отдельные виды имеют обращенный назад (как и первый) второй или же четвертый палец. Следует отметить, что у стрижей первый палец, направлен, как и остальные пальцы, вперед, тогда как у скопы он может поворачиваться в обе стороны. Цевка у птиц на землю не опирается, и они ходят только на пальцах, не опираясь на грунт пяткой.
Мышечная система у птиц
Ноги крылья и прочие часть тела птицы, приводятся в движение с помощью примерно 175 различных скелетных попречнополосатых мышц. Эти мышцы еще называют произвольными, поскольку их сокращения могут контролироваться сознанием и, соответственно, они могут быть произвольными. Как правило, эти мышцы парные, расположенные симметрично на правой и левой стороне тела.
Основными мышцами обеспечивающими полет являются грудная мышца и надкоракоидная. И та и другая мышца начинаются на грудине. Наиболее крупная мышца – грудная. Она тянет крыло вниз обусловливая самым движение птицы в воздухе вверх и вперед. А надкоракоидная мышца поднимает крыло вверх, в противоположном работе грудной мышцы направлении, подготавливая его к очередному взмаху. Надо сказать, что у индейки и домашней курицы, эти две мышцы считаются «белым мясом», тогда как остальные мышцы относятся к «темному мясу».
 Мышечная система птиц.
Мышечная система птиц.Кроме скелетной произвольной мускулатуры, птицы имеют, как и прочие позвоночные, гладкую мускулатуру, которая слоями залегает в стенках органов мочеполовой, пищеварительной, сосудистой и дыхательной систем. Кроме этого гладкие мышцы есть и в коже. Именно ими обусловлены движения перьев. Есть гладкая мускулатура и в глазах: благодаря ей обеспечивается фокусировка изображения на сетчатке. Такую мускулатуру в противоположность поперечно-полосчатой именуют непроизвольной мускулатурой, поскольку она работает без волевого контроля.
Нервная система у птиц
Центральная нервная система птиц состоит из спинного и головного мозга, образованных множеством нейронов нервных клеток.
 Нервная система птиц.
Нервная система птиц.Наиболее заметной частью головного мозга у птиц являются большие полушария, которые представляют собой центр, в котором происходит высшая нервная деятельность. Поверхность этих полушарий не имеет ни извилин, ни борозд, типичных для многих млекопитающих, а ее площадь достаточно мала, что совпадает с относительно низко развитым интеллектом основной массы птиц. Внутри больших полушарий размещаются центры координации тех форм активности, которые связаны с инстинктом, включая инстинкты кормежки и пения.
Особый интерес представляет птичий мозжечок, который находиться сразу за большими полушариями, и покрыт извилинами и бороздами. Его большой размер и строение, соответствуют тем сложным задачам, которые связаны с сохранением в воздухе равновесия и координацией множества движений необходимых для полета.
Сердечно-сосудистая система у птиц
По отношению к размерам тела, сердце у птиц, заметно крупнее чем у млекопитающих того же размера. При этом замечено, что чем мельче конкретный вид птиц, тем более крупным будет его сердце (разумеется, относительно размеров ее тела). К примеру, у колибри масса сердца составляет 2,75% от массы всего тела. Это необходимо, чтобы все многолетающие птицы могли обеспечить быструю циркуляцию крови. То же самое относится и к тем видам пернатых, которые обитают на больших высотах или в холодных областях. И, так же как и у млекопитающих животных, у птиц сердце четырехкамерное.
 Строение кровеносной системы птиц.
Строение кровеносной системы птиц.Частота сердечных сокращений находится в зависимости от размеров сердца и самого животного, а также от степени нагрузки. К примеру, частота сердечных сокращений у отдыхающего страуса составляет около 70 уд/мин, тогда как у колибри во время полета она поднимается до 615 уд/мин. При этом, чрезмерный испуг может испугать птицу настолько, что повысившееся давление может привести к тому, что артерии лопаются и птица умирает.
Так же как и млекопитающие, птицы являются теплокровными животными, При этом, диапазон нормальных температур их тел, у них выше, чем у людей и колеблется в диапазоне от 37,7 до 43,5 градусов. Как правило, птичья кровь содержит больше эритроцитов, чем у основной массы млекопитающих. Благодаря этому птичья кровь может перенести больше кислорода за единицу времени, что очень важно для полета.
Дыхательная система у птиц
Почти у всех птиц ноздри ведут в находящиеся у основания клюва носовые полости. Но есть и исключения: олуши, бакланы и некоторые другие виды пернатых не имеют ноздрей и поэтому вынуждены дышать через рот. Попавший в нос или рот воздух перемещается в гортань, за которой начинается трахея.
 Строение дыхательной системы птиц.
Строение дыхательной системы птиц.В отличие от млекопитающих, гортань птиц звуков не производит, будучи лишь клапанным аппаратом, который защищает нижние дыхательные пути от того, чтобы в них попадала вода и пища.
Ближе к легким трахея разделяется на два бронха, которые входят по одному в каждое легкое. В той точке, где они разделяются, располагается нижняя гортань, которая и служит птице голосовым аппаратом. Образована она окостеневшими расширенными костями трахеи и бронхов, а также внутренними перепонками. К оным прикрепляются пары специальных певчих мышц. Когда выдыхаемый из легких воздух проходит через нижнюю гортань, он приводит к вибрации перепонок, что и производит звуки. У тех птиц, для которых характерен широкий диапазон издаваемых тонов, имеется больше напрягающих голосовые перепонки певчих мышц, чем у тех видов, которые поют откровенно плохо.
Каждый бронх разделяется при входе в легкие на тонкие трубочки. Стенки этих трубочек пронизаны кровеносными капиллярами, которые получают кислород из воздуха и отдают обратно в него углекислый газ. Эти трубочки направляются в тонкостенные воздушные мешки, напоминающие не пронизанные капиллярами мыльные пузыри. Находятся эти мешки за пределами легких – в районе таза, плеч, шеи, вокруг пищеварительных органов и нижней гортани и даже приникают в крупные кости крыльев и ног.
 Птицы способны летать благодаря устройству крыльев и наличию воздушных мешков.
Птицы способны летать благодаря устройству крыльев и наличию воздушных мешков.Когда птица делает вдох, воздух через трубочки попадает в эти самые мешки, а при выдохе он из мешков идет по трубочкам через легкие, где снова происходит газообмен. Благодаря такому двойному дыханию, снабжение организма кислородом увеличивается, что создает более благоприятные условия для полета.
Кроме этого воздушные мешки увлажняют воздух, а также регулируют температуру тела. Это достигается за счет того, что в результате испарения и излучения, окружающие ткани могут терять тепло. В итоге птицы обретают способность, словно потеть изнутри, что является достойной компенсацией отсутствия у птиц потовых желез. Кроме того, воздушные мешки способствуют удалению из организма излишней жидкости.
Устройство пищеварительной системы у птиц
В целом можно сказать, что пищеварительная система птиц представляет собой простирающуюся от клюва вплоть до отверстия клоаки полую трубку. Эта трубка выполняет сразу множество функций, принимая в себя корм, выделяя соки с ферментами, которые расщепляют пищу, всасывает вещества, а также выводит наружу не переварившиеся остатки пищи. Однако, несмотря на то, что у всех птиц строение пищеварительной системы, как и ее функции, одинаковы, в некоторых деталях имеются различия, которые связаны с кормовыми повадками, а также с рационом конкретной группы пернатых.
 Строение пищеварительной системы птиц.
Строение пищеварительной системы птиц.Процесс пищеварения начинается с попадания пищи в рот. Основная масса птиц обладает слюнными железами, которые выделяют смачивающую корм слюну, с него и начинается переваривание пищи. У некоторых птиц, например у стрижеобразных, слюнные железы секретируют липкую жидкость, которая используется для строительства гнезд.
Функции и форма языка, впрочем как и клюва птицы, зависят от того какой образ жизни ведет тот или иной вид птиц. Язык может использоваться как для держания корма во рту, так и для манипуляций с ним в ротовой полости, а также для определения вкуса пищи и ее ощупывания.
Колибри и дятлы, обладают очень длинным языком, который они могут высовывать далеко за пределы своего клюва. У отдельных дятлов на конце языка имеются направленные назад зазубрины, благодаря которым птица может вытащить на поверхность находящихся в коре насекомых и их личинок. А вот у колибри язык, как правило, на конце раздвоен и свернут в трубочку, что помогает высасывать нектар из цветков.
 При помощи языка колибри достает сладкий нектар из цветов.
При помощи языка колибри достает сладкий нектар из цветов.Голубей, фазановых, тетеревиных и индеек, а также у некоторых других птиц, часть пищевода постоянно расширена (ее называют зобом) и используется для накопления пищи. У многих птиц пищевод является достаточно растяжимым и может в течение некоторого времени вмещать в себя значительное количество пищи еще до того как она поступит в желудок.
Желудок у птиц делится на железистую и мускульную («пупок») часть. Железистая часть секретирует, расщепляющий пищу на пригодные для последующего всасывания вещества, желудочный сок. Мускульная часть желудка характеризуется толстыми стенками и твердыми внутренними гребнями, размалывающими пищу, которая получена из железистого желудка, что выполняет компенсирующую функцию для этих не имеющих зубов животных. Особенно толстыми мышечные стенки являются у тех птиц, которые питаются семенами и прочими твердыми кормами. Поскольку часть попавшего в желудок корма может быть непереваренной (например, твердые части насекомых, волосы, перья, части костей и пр.) то у многих хищных птиц в «пупке» образуются округлые плоские погадки, которые время от времени отрыгиваются.
 Благодаря слаженной работе пищеварительной системы, маленькие птенцы вырастают и становятся красивыми птицами.
Благодаря слаженной работе пищеварительной системы, маленькие птенцы вырастают и становятся красивыми птицами.Продолжается пищеварительный тракт тонкой кишкой, которая следует сразу же за желудком. Именно там происходит окончательное переваривание пищи. Толстая кишка у птиц, представляет собой ведущую в клоаку толстую прямую трубку. Кроме нее в клоаку открываются еще и протоки мочеполовой системы. В результате в клоаку попадают как фекальные массы, так и сперма, яйца и моча. И все эти продукты покидают тело птицы через это одно отверстие.
Мочеполовая система у птиц
Мочеполовой комплекс состоит из выделительной и репродуктивной систем, которые очень тесно между собой связаны. Выделительная система функционирует беспрерывно, тогда как вторая активизируется только в определенное время года.
 Мочеполовая система птиц.
Мочеполовая система птиц.Выделительная система состоит из ряда органов, среди которых, в первую очередь, следует назвать две почки, которые извлекают из крови отходы жизнедеятельности и образуют мочу. Мочевого пузыря у птиц нет, поэтому моча через мочеточники поступает прямо в клоаку, где основная масса воды снова всасывается в организм. Оставшийся после этого похожий на кашу белый остаток, вместе с поступившими из толстой кишки фекалиями темного цвета выбрасывается наружу.
Репродуктивная система у птиц
Эта система состоит из половых желез (гонад) и трубок, которые отходят от них. Мужские гонады представлены парой семенников, в которых формируются гаметы (мужские половые клетки) – сперматозоиды. Форма у семенников либо эллиптическая, либо овальная, при этом левый семенник, как правило, крупнее правого. Семенники находится в полости тела рядом с передним концом каждой почки. С приближением брачного сезона гормоны гипофиза, за счет своего стимулирующего действия, увеличивают семенники в несколько сотен раз. По тонкому и извилистому, семявыносящему протоку сперматозоиды из каждого семенника попадают в семенной пузырек. Именно там они и накапливаются, сохраняясь до копуляции и происходящего в этот момент семяизвержения. При этом они попадают в клоаку и выходят наружу через ее отверстие.
 Репродуктивная система птиц.
Репродуктивная система птиц.Яичники (женские гонады) образуют яйцеклетки (женские гаметы). Основная масса имеет только один (левый) яичник. Яйцеклетка, если сравнивать ее с микроскопическим сперматозоидом имеет огромный размер. В плане массы, ее основную часть составляет желток, который является питательным материалом для, начавшего развиваться после оплодотворения, зародыша. Яйцеклетка из яичника попадает в яйцевод, мышцы которого проталкивают яйцеклетку мимо всевозможных железистых областей находящихся в стенах яйцевода. С их помощью желток окружается белком, находящимися под скорлупой оболочками и состоящей по большей части из кальция скорлупой. В конце добавляются пигменты, окрашивающие скорлупу в тот или иной цвет. На то, чтобы яйцеклетка развилась готовое к откладке яйцо, требуется около суток.
Для птиц характерно внутреннее оплодотворение. Во время копуляции сперматозоиды попадают в клоаку самки и затем перемещаются вверх по яйцеводу. Женская и мужская гаметы (то есть, собственно оплодотворение) происходит в верхнем конце яйцевода еще до того, как яйцеклетка будет покрыта белком, подскорлуповыми оболочками и скорлупой.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Мозговой отдел черепа птиц (Aves)
Череп
птиц имеет большие орбиты и тонкую
межглазничную перегородку, сильно
вздутую черепную коробку позади орбит
и вытянутую переднюю часть в клюв.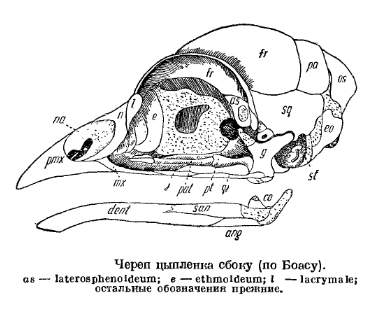
В затылочной области развиваются все четыре затылочные кости. Имеется 1 затылочный мыщелок, лежит на нижней стороне позвоночника. В связи с разрастанием крыши черепа, слуховые капсулы сдвинуты вниз. В слуховой капсуле 3 ушные кости, которые срастаются в одну общую массу. В глазнично-височной области основная и передняя клиновидные кости, а в глазницах парные боковые клиновидные и глазнично-клиновидные кости. В передней части межглазничной перегородки лежит обонятельная кость. Под дном черепа развивается рудиментарный парасфеноид, который срастается с базисфеноидом, сошник развивается как парная кость, но затем срастается.
Мозговой отдел черепа млекопитающих(Mammalia)
Череп
имеет широкое основание, однако это
результат изменения тропибазального
черепа, обусловленный разрастанием
мозговой коробки. У эмбрионов иногда
наблюдаются остатки межглазничной
перегородки.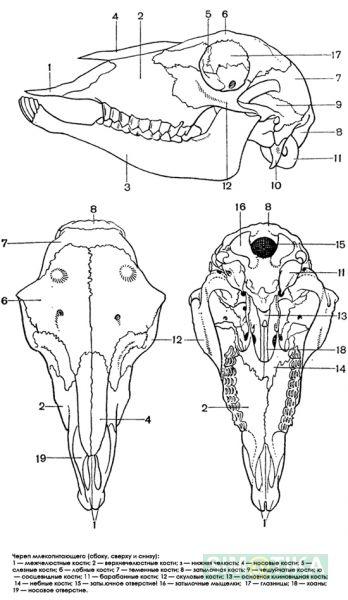
Хрящевой череп эмбрионально иногда представлен довольно полно. Почти сплошной хрящ наблюдается в стенке ушных и обонятельных капсул, в основании и на боках черепа.
В костном черепе наблюдается весьма характерный процесс сокращения числа костей путем их срастания.
В затылочной области имеется 2 затылочных мыщелка. Большое затылочное отверстие ограничена основной и боковыми затылочными костями. Почти всегда все 4 затылочные кости срастаются в одну кость. В слуховой капсуле развивается несколько окостенений, из которых главное сравнивают с предушной, а заднее с заднеушной костью. Эти окостенения очень рано срастаются в каменистую кость (petrosum). У млекопитающих появляется барабанная кость(tympanicum), ограничивающая снаружи полость среднего уха. Барабанная кость вместе с височной срастаются в височную(temporale). В глазнично-височной области развиваются две непарные кости – основная клиновидная и передняя клиновидная. Закладывается рудиментарный парасфеноид. К основной клиновидной кости прирастают височные крылья.
Впереди клиновидных костей в обонятельной области из нескольких окостенений развивается решетчатая кость (etmoideum). Отдельно окостеневает средняя часть, образуя носовую перегородку.
Крыша черепа образована теменными и лобными костями. Теменные кости нередко срастаются, между ними появилась еще одна кость – межтеменная(interparietale). У переднего края глазницы есть небольшая слезная кость(lacrimale).
Височная область черепа прикрыта скуловой дугой, составленной из скуловой кости и скуловых отростков чешуйчатой и челюстной костей.
Вторичные верхние челюсти образуют переднюю нижнюю часть черепа. У прдчелюстных и челюстных костей отсутствует отросток, ограничивающего ноздри.
Твердое небо образовано за счет небных отростков межчелюстных, челюстных и небных костей, которые сходятся на средней линии и образуют шов. Небные кости, как и крыловидные, стоят вертикально и ограничивают хоаны с боков. Крыловидные кости вполне самостоятельны.
Кольцо с птичьим черепом, похоже на настоящий череп, но вырезано из кости, поэтому оно красивое и крепкое. Кость становится белой, но я впитываю ее в чай, чтобы придать ей эффект старины и выделить резные детали. Если вы хотите, чтобы он был темнее, вы можете замочить его в крепком кофе.
Кончик клюва может быть острым. Некоторые люди любят это (подсказка, это отлично подходит для чистки ногтей или царапин вещей).) Если вы хотите, чтобы ваш наконечник был мягче, аккуратно напишите наждачной доской, пока он не станет правильным.
Если вы не видите предложенного вами размера, я рекомендую увеличить его вдвое или даже полностью. Вы всегда можете вставить небольшую кольцевую ленту под клювом для дополнительной безопасности.
Это поразительная вещь и веселый стартер разговора.
Идеально подходит для раздачи конфет детям на Хэллоуин.
Это выглядит и чувствует себя хорошо с клювом, указывающим по Вашей руке, или по Вашему пальцу. Это удобно и сильно, но я все равно рекомендовал бы избегать ударов при ношении.Не нужно играть в боулинг, бонго или викингов, но набирать тексты очень весело, потому что кажется, что они клюют клавиши. Я также постараюсь свести к минимуму воздействие воды, так как со временем хлор и т. Д. Может высушить кость. Вся кость – это коровья кость, этически полученная, только остатки тех, кто решил есть мясо.
Более подробную информацию о моих ювелирных украшениях и соответствующих серьгах и ожерельях можно найти здесь:
www.etsy.com/shop/TalismanaDesigns?ref=seller-platform-mcnav§ion_id=14296529
Я ЛЮБЛЮ быструю доставку и обычно высылаю в течение 24 часов Получив ваш заказ:)
Для хорошего времени посетите меня на Facebook: www.facebook.com/talismanadesigns или instagram
Дарите лайк на моей странице и сделайте мой день; )
~ См. Раздел «Правила магазина» для получения полной информации, вот несколько важных битов:
– Возврат или обмен – Пожалуйста, отправьте обратно в течение 14 дней с момента получения вашего товара. Он должен быть на 100% не изношенным и в повторно продаваемом состоянии. Пожалуйста, позаботьтесь о том, чтобы на нем не было шерсти домашних животных.
– Любая бесплатная доставка с 10% комиссией за возврат.
– Международные покупатели несут ответственность за любые пошлины, взимаемые их страной.
– Etsy автоматически отправит вам информацию об отслеживании, как только я распечатаю ваш ярлык с доставкой,
– Если ваша посылка доставлена, но отсутствует в вашей почте, пожалуйста, свяжитесь с семьей, соседями по комнате и соседями, а также с местным почтовым отделением или почтой перевозчик. И проверьте глубину почтового ящика, так как некоторые пропущенные пакеты, очевидно, прячутся там.
– рекомендуется стирка в холодной воде, деликатный цикл, дать высохнуть на воздухе. Предметы, окрашенные вручную, следует стирать отдельно с первого раза.
Спасибо за помощь в развитии моих творческих замыслов!
,Etsy использует файлы cookie и аналогичные технологии, чтобы обеспечить вам лучший опыт, обеспечивая такие вещи, как:
- Основные функции сайта
- обеспечение безопасных, безопасных транзакций
- безопасный вход в аккаунт
- запоминание учетной записи, браузера и региональных предпочтений
- запоминание настроек конфиденциальности и безопасности
- анализ трафика и использования сайта
- персонализированный поиск, содержание и рекомендации
- помогает продавцам понять свою аудиторию
- показывает релевантные целевые объявления на Etsy и за его пределами
Подробную информацию можно найти в Политике Etsy в отношении файлов cookie и аналогичных технологий и в нашей Политике конфиденциальности.
Всегда на
Требуемые файлы cookie и технологии
Некоторые из технологий, которые мы используем, необходимы для критически важных функций, таких как безопасность и целостность сайта, аутентификация учетной записи, настройки безопасности и конфиденциальности, данные внутреннего использования и обслуживания сайта, а также для правильной работы сайта для просмотра и транзакций.
отНастройка сайта
Файлы cookie и аналогичные технологии используются для улучшения вашего опыта, например:
- запомнить ваш логин, общие и региональные предпочтения
- персонализировать контент, поиск, рекомендации и предложения
Без этих технологий такие вещи, как персональные рекомендации, настройки вашей учетной записи или локализация, могут работать неправильно.Узнайте больше в нашей Политике в отношении файлов cookie и аналогичных технологий.
отПерсонализированная реклама
Эти технологии используются для таких вещей, как:
- персонализированных объявлений
- , чтобы ограничить количество просмотров рекламы .
- , чтобы понять использование через Google Analytics
- чтобы понять как ты добрался до Etsy
- , чтобы убедиться, что продавцы понимают свою аудиторию и могут предоставить релевантную рекламу
Мы делаем это с партнерами по социальным сетям, маркетингу и аналитике (у которых может быть своя собственная информация, которую они собрали).Сказать «нет» не помешает вам видеть рекламу Etsy, но это может сделать их менее актуальными или более повторяющимися. Узнайте больше в нашей Политике в отношении файлов cookie и аналогичных технологий.
,